Majungatholus atopus: Etymology: The generic name is derived from the name of the region where the specimen was found and the Latin tholus, 'dome'.The specific epithet is derived from the Greek atopos, 'strange' (or, in its original derivation, 'out of place' — a perhaps more appropriate meaning in the present context).
Diagnosis: A single dome-like thickening of the frontal region; frontal dome thick, with highly irregular, rugose dorsal surface. Parietal without significant thickening, participating in a well developed parieto-squamosal shelf. Very large supratem-poral fenestrae. Olfactory portion of the braincase long; olfactory lobes ventrally enclosed by bone (Sues and Taquet, 1979). Medium-sized pachycephalo-saurid with single well-developed dome formed by fused and thickened frontals. Dorsal surface of dome with highly irregular ornament of nodes and furrows; median depres sion on parietals without significant thickening, participating in formation of parietosquamosal shelf. Supratemporal fenestrae large. Olfactory stalks very long; olfactory lobes almost completely enclosed by bone ventrally (Sues, 1980). Diagnostic characters for Majungatholus atopus: thickened, fused, highly pneumatic nasals with large, bilateral foramina; frontals with median hornlike projection; cervical ribs bifurcate distally; cervical ribs with multiple enlarged pneumatic foraminae proximally (diameter . 10 mm) (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998).
Holotype: Partial skull, consisting of the frontals and parietals (the latter being somewhat damaged posteriorly) and the anterior part of the braincase. Formerly in the collections of the Ecole des Mines, Paris (uncata-logued), now in the collections of the Museum National d'Histoire Naturelle, Paris, MNHN.MAJ 4. (formerly in the collections of the Ecole des Mines, Paris).
The specimen, a detailed anatomical description of which will be presented elsewhere, evidently represents a dome-headed dinosaur referable to the Pachycephalosauridae. Like other pachycephalosaurids, it differs from the Ankylosauria in showing very long olfactory stalks and thickening of the cranial roof bones by upgrowth of the bones rather than by fusion of dermal ectopic elements. Furthermore, in ankylosaurs the upper temporal fenestrae are always closed. Majungatholus represents a distinctive type of the dome-headed ornithischians; its single frontal dome and its very large upper temporal fenestrae are unique features. The earliest known pachycephalosaurid, Yaverlandia bitholus, from the Lower Cretaceous (Wealden) of the Isle of Wight 1 , shows some resemblance in having large supratemporal fenestrae and thickening restricted to the frontal region; it differs, however, in having a small dome on each frontal and in being much smaller. The only Upper Cretaceous pachycephalosaurid species showing a certain similarity is a flat-headed form referred to Stegoceras from the Campanian of Alberta, Canada . In this species the supratemporal fenestrae are large and a parieto-squamosal shelf is well developed; the frontals, however, are but slightly thickened and the species is smaller than Majungatholus atopus. A peculiar feature only present in Majungatholus and the two Laurasian species mentioned above is a median circular depression on the postero-dorsal surface of the frontals. Majungatholus can be derived from a Yaverlandia-\ike form and probably represents a lineage evolving in geographical separation from Northern Hemisphere pachycephalosaurids. The presence of a pachycephalosaurid in Madagascar and of other 'Laurasian' dinosaur families in the Southern Hemisphere during the Late Cretaceous can be explained in two different ways. First, dispersal from Laurasia could be assumed as all these families have their greatest diversity and most primitive species in the Northern Hemisphere. The Tethyan sea belt separated the two super-continents during the Cretaceous but it was at least in part a fairly shallow epicontinental sea and passage routes may have become available on several occasions. The exact locations of some of these connections may never be known as many potential areas have subsequently been affected by Cretaceous and Tertiary orogeneses. Cox has suggested the existence of a filter route in the Central American or Carribean region to account for the presence of a hadrosaur in Argentina . But it can also be argued that the existence of similar dinosaurs in both hemispheres is the result of vicariance events (the breakup of Pangea). The evidence presently available does not permit a choice between the two hypotheses. It is clear that faunal exchange by land connections between Laurasia and Gondwanaland was possible at least during the first half of the Cretaceous, possibly even at a later date (Sues and Taquet, 1979).
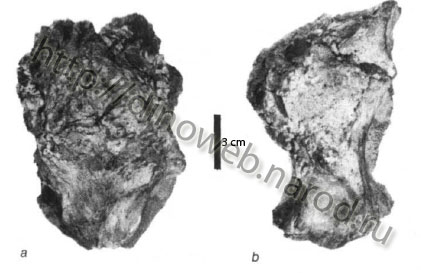
Majungatholus atopus,MNHN.MAJ 4. Partial skull in dorsal (a) and lateral (b) view (modified from Sues and Taquet, 1979).
|  DINOWEB - палеозойские и мезозойские тетраподы
DINOWEB - палеозойские и мезозойские тетраподы