Majungasaurus
Genus: Majungasaurus Lavocat, 1955
Classification: Dinosauria - Saurischia - Theropoda - Ceratosauria - Abelisauroidea - Abelisauridae - Carnotaurinae
Etymology: From Majunga, in reference to the province of Mahajanga (formerly Majunga), and saurus, New Latin, "lizard”, from the Greek sauros.
Synonyms: = Majungatholus Sues and Taquet, 1979
Type species: M. crenatissimus (Deperet, 1896) Lavocat, 1955
Other Species: none
Diagnosis: as for type species
Species: Majungasaurus crenatissimus (Deperet, 1896) Lavocat, 1955
Etymology: From crenatus, Latin, "notched or toothed”, and issimus, "most or very much”; in reference to "the serrations which are extended along the entire length of the two trenchant ridges of the teeth” (Deperet, 1896).
Synonyms: = Megalosaurus crenatissimus Daperet, 1896; = Dryptosaurus crenatissimus Deperet and Savornin;, 1928= Majungatholus atopus Sues and Taquet, 1979.
Holotype: MNHN.MAJ 1, fragmentary right dentary of a subadult individual (Lavocat, 1955).
Referred Specimens: MNHN.MAJ 4 (holotype of Majungatholus atopus) lacrimal fragment, frontals, parietals, mesethmoid, laterosphenoids. FMNH PR 2008, right premaxilla. This specimen is provisionally assigned to M. crenatissimus on the following grounds: 1) serration morphology on the premaxillary teeth is indistinguishable from that present on the two teeth present in the original collection described by Deperet (1896), as well as on hundreds of isolated teeth found in Upper Cretaceous deposits (Maevarano Formation) of the Mahajanga Basin in 1993 and 1995; 2) among the hundreds of isolated teeth collected in the field area, there is no evidence for more than one large theropod taxon. Exact locality data are unknown for the M. crenatissimus specimens described by Deperet (1896) and Lavocat (1955), though all were found in the Mahajanga Basin, northwestern Madagascar, southeast of Mahajanga. The premaxilla described below (FMNH PR 2008) was recovered from the same field area, at a locality designated MAD93-33. The geographic coordinates of MAD93-33, established with a satellite Global Positioning System (GPS), are: latitude 15°, 54", 28.31' S and longitude 46°, 34", 56.38' E. FMNH PR 2008–right premaxilla; F MNH PR 2099–partial skull roof including partially fused frontals with small median cornual process, or ‘horncore,’ of immature individual ; FMNH PR 2100–nearly complete, exquisitely preserved, disarticulated skull (missing only left premaxilla, right pterygoid, right ectopterygoid, both epipterygoids, both vomers, and both columellae), with 26 associated caudal vertebrae and 18 haemal arches; FMNH PR 2278–associated cranial (both premaxillae, both maxillae, left jugal, left quadratojugal, left ectopterygoid, left quadrate, left surangular, left angular, left prearticular, and left articular), and postcranial (two fragmentary cervical neural arches, three fragmentary dorsal vertebrae, one fragmentary caudal vertebra, left scapulocoracoid, partial left ilium, left femur, left and fragmentary right tibiae, left and partial right fibulae, left astragalocalcaneum, left metatarsals II-IV, right pedal phalanges II-1 and IV-2, and left pedal phalanges IV-2 and IV-3) elements of large near-adult; FMNH PR 2293–axis (C2) vertebra; FMNH PR 2294–five articulated caudal vertebrae (the last three fused pathologically) with two haemal arches; FMNH PR 2295–3rd cervical vertebra (C3); FMNH PR 2423–right humerus; FMNH PR 2424–left tibia; FMNH PR 2425–left astragalocalcaneum; FMNH PR 2426–right pedal phalanx II-1; FMNH PR 2427–right pedal phalanx II-2; FMNH PR 2428–left pedal phalanges II-3 and III-2 and the distal end of right pedal phalanx II-1; FMNH PR 2429–left pedal phalanx III-1; FMNH PR 2430–right pedal phalanx IV-1; FMNH PR 2431–left pedal phalanx IV-3; FMNH PR 2432–left pedal phalanx IV-4; FMNH PR 2433–right pedal phalanx IV-4; FMNH PR 2434–left pedal phalanx IV-5; FSL 92.290–pedal ungual phalanx; MNHN.MAJ 4–partial skull roof with portions of partially fused frontals (with rounded median cornual process), parietals, caudal process of right lacrimal, sphenethmoid, and laterosphenoids; UA 8678–incomplete and disarticulated skull (including left splenial, left prearticular, right surangular, and right squamosal) of subadult individual associated with 23 presacral vertebrae, 13 cervical and 14 dorsal ribs, a partial sacrum, five proximal caudal vertebrae, one middle caudal vertebra, the first haemal arch, and both ilia; UA 8709–nearly complete, articulated, but poorly preserved skull (including maxillae, nasals, frontals, jugals, lacrimals, right postorbital and squamosal, pterygoids, ectopterygoids, right palatine, and partial braincase) and both lower jaws; UA 8716–right premaxilla; UA 8717–right and left premaxillae; UA 8718–partial left lacrimal; UA 8719–partial skull roof; UA 8782–distal portion of left quadrate; UA 9031–left humerus; UA 9032–left tibia; UA 9033–right astragalocalcaneum; UA 9034–left metatarsal II; UA 9035–left metatarsal IV; UA 9036–left pedal phalanx II-1; UA 9037–right pedal phalanx II-2; UA 9038–left pedal phalanx II-3; UA 9039–right pedal phalanx III-1; UA 9040– right pedal phalanx IV-1; UA 9041–right pedal phalanx IV-2; UA 9042–left pedal phalanx III-2; UA 9043–pedal phalanx IV-5; UA 9077–left tibia and fibula; UA 9078– right fibula; UA 9079–left metatarsal III; UA 9081–right pedal phalanx III-1 or III-2; UA 9082–astragalus; UA 9089– four proximal caudal vertebrae; UA Bv 532–left pedal phalanx I-2; UA Bv 1658–left pedal phalanx I-2; UA Bv 1260–right pedal phalanx II-1; UA Bv 1265–left pedal phalanx III-1; plus literally thousands of isolated teeth (including FMNH PR 2198 and 2228, FSL 92.306a-b, MNHN 1911a-d, MNHN.MAJ 242, MNHN.MAJ 249) from numerous localities in the Berivotra Study Area. MSNM V5255, MSNM V5256, MSNM V5267, MSNM V5276, MSNM V5509, MSNM V5510, MSNM V6418, MSNM V6419, MSNM V6420, MSNM V6421, pedal elements.
Time: Late Cretaceous (Maastrichtian)
Horizon: Maevarano Formation
Location: Mahajanga Basin, northwestern Madagascar, in the small field area surrounding the village of Berivotra, southeast of the port city of Mahajanga.
Total length: 7-9 m.
Mass: 1-2 tonns.
Diagnosis: Abelisaurid theropod with: (1) dorsoventrally deep, fused nasals that are strongly pneumatized via large, bilateral foramina; (2) nasal processes of left and right premaxillae separated by a thin lamina of nasal; (3) maxilla bearing 17 alveoli; (4) frontals rounded rostrally rather than forming a double notch; (5) frontals with a sculptured, median cornual process in adults that is variably pneumatic; (6) pronounced median fossa on sagittal (frontoparietal) crest; (7) dentary bearing 17 alveoli, virtually no extension caudal to last alveolus, and ventral position of lateral sulcus; (8) teeth bearing weakly developed interdenticular sulci; (9) long, falciform atlantal epipophysis; (10) vertebral centra in cranial dorsal series with dorsoventrally elongate articular surfaces; (11) cranial and caudal borders of midcervical transverse processes parallel in lateral view; (12) dorsal and caudal vertebral neural spines dorsally expanded (transversely and craniocaudally); and (13) cervical ribs pneumatized via multiple, enlarged foramina on medial surface of shaft, and accessory foramina on cranial and caudal surfaces of capitulotubercular web (Krause, Sampson, Carrano and O'Connor, 2007).
Comments:
About a century ago, French paleontologist Charles Deperet received a shipment of vertebrate fossils recovered from Upper Cretaceous strata near the town of Mahajanga (=Majunga), northwestern Madagascar. He recognized six specimens as the remains of a large theropod, and placed them in a new species of Megalosaurus, M. crenatissimus (Deperet, 1896). Lavocat (1955) later assigned an incomplete dentary, also recovered from the Mahajanga Basin, to this species, but erected a new genus, Majungasaurus. Russell et al. (1976) reported the discovery of a partial skeleton of M. crenatissimus from the Mahajanga Basin, but this specimen remains unprepared and unstudied in the Service de Paleontologic, Universite d' Antananarivo. Additional Late Cretaceous theropod remains purported to resemble those of M. crenatissimus include isolated teeth and terminal phalanges from Egypt (Gemellaro, 1921; Stromer and Weiler, 1930), isolated teeth from Argentina (Bonaparte and Powell, 1980), and an incomplete tooth from India (Mathur and Srivastava, 1987). While some workers have considered Majungasaurus a "megalosaurid" (e.g., Molnar, 1980), others have suggested tyrannosaurid affinities (e.g., Charig, 1973; Sues, 1980). Sues (1980: 959) concluded with regard to M. crenatissimus that the "scanty remains ... are too fragmentary to allow assessment of the affinities of the species." Bonaparte (1986, 1991) referred Majungasaurus "with doubts" to Abelisauridae. Molnar (1990) tentatively supported the abelisaurid designation, noting that Carnotaurus sastrei from the Middle Cretaceous of Argentina possesses a similarly curved dentary (Bonaparte et al., 1990), yet relegated Majungasaurus to Theropoda incertae sedis. In the austral winter of 1993, a joint SUNY-Stony Brook! Universite d' Antananarivo expedition recovered numerous specimens of fossil vertebrates from Upper Cretaceous terrestrial deposits in the Mahajanga Basin, northwestern Madagascar (Fig. 1; Krause and Hartman, 1996). Among these was a nearly complete, exquisitely preserved premaxilla of the poorly known large theropod, Majungasaurus crenatissimus. This element, the first known cranial remains of M. crenatissimus, possesses derived morphology supporting placement of the taxon within the ceratosaurian clade Abelisauridae. It also shares several synapomorphies with the premaxilla of Indosuchus raptorius, a large theropod from the Late Cretaceous of India. Other than overall size, there are no significant differences in the premaxillae of these two taxa, supporting contentions that: 1) Majungasaurus and Indosuchus are sister taxa within Abelisauridae; and 2) Madagascar and the Indian subcontinent remained part of the same biogeographic province into the Late Cretaceous (Sampson, Krause, Dodson and Forster, 1996).
Premaxilla of Majungasaurus crenatissimus in lateral (A) and medial (B) views (modified from Sampson, Krause, Dodson and Forster, 1996). Recent discoveries of fossil vertebrates from the Late Cretaceous of Madagascar include several specimens of a large theropod dinosaur. One specimen includes a nearly complete and exquisitely preserved skull with thickened pneumatic nasals, a median frontal horn, and a dorsal projection on the parietals. The new materials are assigned to the enigmatic theropod group Abelisauridae on the basis of a number of unique features. Fossil remains attributable to abelisaurids are restricted to three Gondwanan landmasses: South America, Madagascar, and the Indian subcontinent. This distribution is consistent with a revised paleogeographic reconstruction that posits prolonged links between these landmasses (via Antarctica), perhaps until late in the Late Cretaceous (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998). One of the theropod specimens (FMNH PR 2100) includes a nearly complete skull —among the best preserved and most complete dinosaur skulls known—and most of the tail. The skull is disarticulated and individual bones are virtually undistorted, allowing comprehensive and detailed study of all elements. The external surface of many elements is covered in rugose sculpturing, and the skull roof is adorned with three median ornamentations: thickened, fused nasals; a low frontal horn; and a parietal eminence. The total skull length is 57 cm, and comparisons with a closely related taxon, Carnotaurus sastrei from Argentina, suggest a total adult body length of about 7 to 9 m. A second specimen (UA 8678) of the same taxon includes an incomplete and disarticulated skull, most of the precaudal axial column, and the left ilium. Several of the vertebrae and ribs, particularly in the cervical region, were recovered in articulation. The small size of the skull elements relative to those of FMNH PR 2100, combined with the lack of fusion between several vertebral centra and corresponding neural arches, indicates that this animal was immature at the time of death. Although large theropod materials from the Maevarano Formation have generally been referred to Majungasaurus crenatissimus, the inadequacy of the holotype and neotype specimens requires that this taxon be regarded as a nomen dubium. Comparison of the recently collected materials with the fragmentary holotype specimen of the putative Malagasy pachycephalosaur, Majungatholus atopus, demonstrates that Majungatholus is not a pachycephalosaur but rather a "domed” theropod. This finding has biogeographic significance in that it removes the only report of a pachycephalosaur from a Gondwanan landmass, thereby restricting occurrences of this dome-headed ornithischian clade to Laurasia. Thus, the materials described herein are referred to Majungatholus atopus and placed within the enigmatic theropod group Abelisauridae (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998).
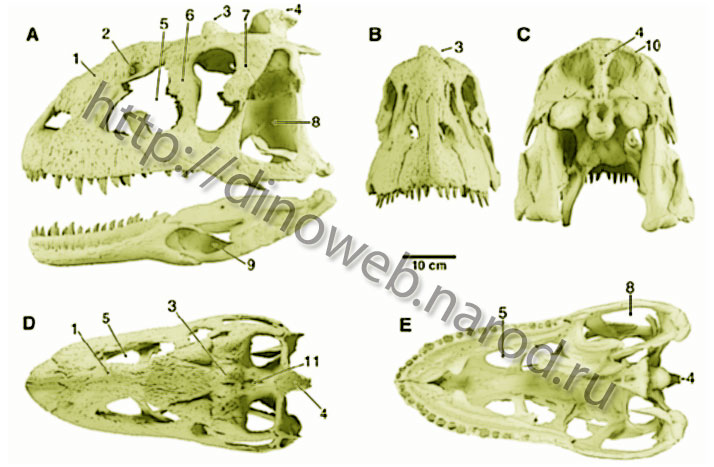
Reconstruction of the skull and lower jaws of Majungatholus atopus, based on preserved elements of FMNH PR 2100 (12). (A) Left lateral view of skull and lower jaws; (B) rostral view of skull; (C) occipital view of skull; (D) dorsal view of skull; (E) palatal view of skull. Numbered features: 1, thickened nasals; 2, pneumatic foramen of nasal; 3, frontal horn; 4, parietal eminence; 5, enlarged antorbital fenestra; 6, lacrimal with suborbital process; 7, postorbital with suborbital process; 8, enlarged laterotemporal fenestra; 9, enlarged external mandibular fenestra; 10, nuchal crest; 11, median depression between frontal horn and parietal eminence (modified from Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998). The known distribution of abelisaurids (Argentina, Indian subcontinent, and Madagascar), like that for gondwanathere mammals, is consistent with at least two major biogeographic hypotheses: Abelisaurids originated before the major continental fragmentations of the Early Cretaceous, and spread throughout most of Gondwana and perhaps into Laurasia. In keeping with this scenario, the current absence of documented African abelisaurids is attributable to poor sampling, differential extinction, or both. Abelisaurids originated sometime in the Early Cretaceous after the tectonic isolation of Africa. If so, abelisaurids never existed on Africa but rather dispersed between South America and Indo-Madagascar via Antarctica, making use of the postulated land bridge across the Kerguelen Plateau. Both views are equally probable given the data at hand. Reconstructions of dinosaur biogeography initially focused on distinctions between Laurasian and Gondwanan faunas. More recently, it has been argued that dinosaur faunas were relatively cosmopolitan until the beginning of the Late Cretaceous, at which time the isolation of continental landmasses by oceanic barriers resulted in an abrupt shift toward marked provincialism. Recent paleogeographic and paleontological data, including those presented here for abelisaurid theropods, suggest an alternative hypothesis: African faunas became increasingly endemic during the Cretaceous after Africa’s isolation from South America, whereas retention of subaerial connections among the remaining Gondwanan landmasses resulted in relatively extensive cosmopolitanism for the associated terrestrial faunas, perhaps until late in the Late Cretaceous (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998). Synapomorphies of Abelisauridae observable in Majungatholus atopus include Cranial: External sculpturing of craniofacial elements; premaxilla with reduced to absent palatal process; maxilla-jugal contact elongate; rostral process of lacrimal strongly reduced or absent; lacrimal with pronounced suborbital process; broad lacrimal-postorbital contact; long axis of postorbital oriented rostroventral-caudodorsal; postorbital with pronounced suborbital process; high, transversely broad nuchal crest with large squamosal contributions; medial alveolar border of premaxilla, maxilla, and dentary bearing fused interdental plates with series of vertical ridges and grooves; hypertrophied external mandibular fenestra and associated weak contacts between dentary and postdentary elements. Postcranial: Postaxial epipophyses elongate (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998). Synapomorphies of Majungatholus and Carnotaurus include Cranial: Postorbital rostrocaudally elongate; postorbital with stepped-down ventrolateral fossa; dorsally placed median fossa in saddle-shaped depression overlapping contact of frontals and parietals; enlarged, caudodorsally projecting parietal eminence supported ventrally by stout supraoccipital; laterally directed paroccipital processes with upturned distal ends; stout, triangular splenial with straight caudal margin) (Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998).
Many lines of evidence have been brought to bear on the question of theropod feeding ecology, including functional and physiological considerations, morphological constraints, taphonomic associations, and telling—although rare—indications of direct ingestion. Tooth marks of theropods, although rarely described and generally left unassigned to a particular taxon, can provide unique clues into predator–prey interaction, and can also yield insights into the extent of carcass utilization. Here we describe a sample of tooth-marked dinosaur bone recovered from three well-documented localities in the Upper Cretaceous Maevarano Formation of Madagascar that provides insights into the feeding ecology of the abelisaurid theropod Majungatholus atopus. Intensely tooth-marked elements from multiple individuals show that Majungatholus defleshed dinosaur carcasses. Furthermore, Majungatholus clearly fed upon the remains of not only sauropods, but also conspecifics, and thus was a cannibal. Cannibalism is a common ecological strategy among extant carnivores, but until now the evidence in relation to carnivorous dinosaurs has been sparse and anecdotal (Rogers, Krause, and Curry Rogers. 2003).
Majungasaurus crenatissimus has had a complex taxonomic history. The original sample, collected from several localities near the Betsiboka River, was described by Deperet (1896a, b) and included two teeth, an ungual phalanx, two partial sacral vertebrae, and a partial caudal vertebra. Deperet named the new species and assigned it to Megalosaurus, but he did not designate any of the six specimens comprising the type series as the holotype and, in addition, a type locality was not specified. Nor did Deperet indicate whether the specimens had been derived from a single individual or, indeed, even the same locality. Deperet and Savornin (1928) later assigned M. crenatissimus to Dryptosaurus, along with D. superbus (=Erectopus), D. aquilunguis, and D. saharicus (=Carcharodontosaurus). Deperet (1896b:191) had earlier revealed his inclination for this assignment by stating the following: "The type species D. aquilunguis Cope is very large, attaining the size of M. bucklandi; the teeth differ entirely from those of Megalosaurus in the serrations of the anterior edge, which are extended along the entire length of this edge to near the root, instead of being limited to the upper part of the crown. It is interesting to note that this character is found exactly in the Malagasy and Indian form, therefore the attribution to the genus Dryptosaurus is shown as entirely probable.” Nearly 60 years after publication on the original sample by Deperet (1896a, b), Lavocat (1955a, b) described a dentary from the same general field area (though several km to the north) and convincingly argued that it represented the same species of theropod as the six specimens described by Deperet. This argument was based primarily on dental morphology because several teeth within and associated with the dentary were regarded as identical to the two teeth described by Deperet. Lavocat, however, recognized that the morphology of the dentary was different from that of other species of Megalosaurus and therefore assigned M. crenatissimus to a new genus, Majungasaurus. Although not explicitly stated, it appears that Lavocat (1955a, b) did not consider any of the specimens in Deperet’s type series to be worthy of designation as the lectotype; instead, he opted to designate the dentary as the neotype of the species. This same specimen, however, was later considered to be nondiagnostic by Sampson and colleagues (1998), and Majungasaurus crenatissimus was thereby regarded as a nomen dubium. In its place, Sampson and colleagues (1998) designated Majungatholus atopus as the valid name for the Maevarano theropod. This followed the 1996 discovery of a theropod skull with lower jaws, which clearly showed that the skull roof (MNHN.MAJ 4) identified by Sues and Taquet (1979) as representing a new genus and species of pachycephalosaur, Majungatholus atopus, instead belonged to a theropod. Sampson and colleagues (1998) argued that, because Deperet did not explicitly designate a holotype for the mid-sized Malagasy theropod, and because Lavocat’s dentary (MNHN.MAJ 1) was non-diagnostic, Majungatholus atopus must be the valid name assigned to the mid-sized theropod from the Maevarano Formation. The recovery and analysis of additional dentary material and especially dental morphology, along with detailed comparison to the dentary (MNHN.MAJ 1) and teeth described by Lavocat (1955), compel us to once again reconsider the taxonomic designation of this theropod. The dentary described by Lavocat (1955), MNHN.MAJ 1, is typically abelisaurid in being broad, laterally bowed (in dorsal view), and relatively foreshortened, as well as in possessing a much-enlarged external mandibular fenestra (resulting in a relatively short post-alveolar portion, terminating caudally slightly behind the last alveolus), pronounced longitudinal sulcus on the lateral surface containing neurovascularm foramina, and rugose subcutaneous texture on its external surfaces (as on the skull). These same features are found in the dentaries of FMNH PR 2100 and UA 8709. As outlined by Sampson and Witmer the adult dentaries of this species can be distinguished from those of other known abelisaurids (e.g., Carnotaurus and the Indian taxon represented by AMNH 1960 [referred to Indosuchus raptorius by Chatterjee, 1978]) by a suite of features, most notably the presence of 17 alveoli, the virtual lack of an extension of the dentary caudal to the last alveolus, and the ventral position of the lateral sulcus. The subadult dentary,MNHN.MAJ 1, possesses these same features and is therefore distinctive among theropods. Indeed, Sampson and Witmer state that "detailed comparisons of MNHN.MAJ 1 with the dentary of FMNH PR 2100 revealed no notable differences other than size (MNHN.MAJ 1 being approximately 25% smaller).” Furthermore, detailed examination of the teeth preserved in MNHN.MAJ 1 by Smith (this volume) reveals that they fall within the same size and shape parameters as the two isolated teeth (FSL 92.306a-b) described by Deperet (1896a, b), those preserved in the gnathic elements of adult individuals described by Sampson and Witmer ( e.g., FMNH PR 2008, 2100; UA 8709, 8716, and 8717), and the thousands of isolated teeth recovered from the Maevarano Formation by Mahajanga Basin Project personnel and others. The teeth of the Malagasy abelisaurid most closely resemble those of AMNH 1753, 1955, and 1960 from the Late Cretaceous of India (referred to Indosuchus raptorius by Chatterjee, 1978) but, importantly, the Malagasy taxon possesses weakly developed interdenticular sulci whereas the Indian specimens lack them. These sulci can also be seen on the erupting teeth of MNHN.MAJ 1. The possession of distinctive features in the dentary and teeth described by Lavocat (1955) and their full morphological congruence with other specimens attributed to the mid-sized Malagasy taxon from the Maevarano Formation (not to mention the lack of evidence for any other mid-sized theropod in the Maevarano Formation, despite the recovery of thousands of vertebrate specimens), ineluctably leads to the conclusion that MNHN.MAJ 1 represents a subadult individual of the same taxon represented by other dentaries (FMNH PR 2100, UA 8709) and countless isolated teeth. As such, and according to the Principle of Priority of the International Code of Zoological Nomenclature (1999), MNHN.MAJ 1 must be designated as the valid type specimen and Majungasaurus crenatissimus as the valid name of this theropod (Krause, Sampson, Carrano and O'Connor, 2007).
The abelisaurid theropod Majungasaurus crenatissimus inhabited the plains of northwestern Madagascar during the Late Cretaceous. It lived alongside other nonavian dinosaurs, including a small-bodied noasaurid theropod (Masiakasaurus knopfleri) and a titanosaurian sauropod (Rapetosaurus krausei). Although an inhabitant of the expansive floodplains of the Mahajanga Basin, M. crenatissimus also frequented the broad and sandy channel belts that drained Madagascar’s central highlands. These shallow rivers were populated by a variety of aquatic and semi-aquatic animals, including fish, frogs, turtles, and several species of both large and small crocodyliforms. These animals were likely adapted for seasonal fluctuations in water availability because the sediments that entomb their remains (fine-grained debris flow deposits intercalated with stream flow deposits) indicate a strongly variable discharge regime. Associated oxidized calcareous paleosols with localized accumulations of carbonate nodules suggest that the ambient climate was semiarid. The numerous bonebeds preserved in these same sediments are indicative of localized and recurrent pulses of mortality. M. crenatissimus fed on carcasses preserved in these bonebeds, and there is good indication from a wealth of toothmarked bone derived from two conspecific individuals that it focused on the well-muscled axial skeleton in a fashion similar to that of many modern vertebrate carnivores. This evidence for intraspecific feeding renders M. crenatissimus the only theropod dinosaur with demonstrated cannibalistic tendencies (Rogers, Krause, Curry Rogers, Rasoamiaramanana and Rahanarisoa, 2007).
Recent fieldwork in the Upper Cretaceous (Maastrichtian) Maevarano Formation, northwest Madagascar, has yielded important new skull material of the abelisaurid theropod, Majungasaurus crenatissimus. One of these specimens in particular—a virtually complete, disarticulated, and well preserved skull—greatly elucidates the craniofacial osteology of abelisaurids. Herein we describe the skull and lower jaws of this mid-sized theropod dinosaur. A number of features of the facial skeleton and cranium (as well as the postcranium) appear to result from increased levels of mineralization and ossification, which, at least in some instances, can be related directly to specific soft-tissue structures; examples include lacrimal-postorbital contact dorsal to the orbit, suborbital processes of the lacrimal and postorbital, presence of a mineralized interorbital septum, fused interdental plates, and mineralization of the overlying integument. Autapomorphic features include a highly derived nasal—greatly thickened and fused to its counterpart, with a large interior pneumatic chamber—and a median, ‘dome'-like thickening of the frontals, which appear to have been variably pneumatized by a paranasal air sac. Majungasaurus also possesses a derived suite of skull morphologies, including: a rostrocaudally abbreviated, dorsoventrally deep, and transversely broad skull; an expanded occiput, likely associated with expanded cervical musculature; short-crowned dentition; and an enlarged external mandibular fenestra consistent with a moderate degree of intramandibular movement or accommodation. A number of characters, present on both the skull and postcranial skeleton, suggest a divergent mode of predation relative to other, non-abelisaurid theropods ( Sampson and Witmer, 2007).
Despite the known diversity of abelisaurid theropod dinosaurs, their dental anatomy remains poorly understood. Discoveries of elements preserving in situ dentition of the Late Cretaceous (Maastrichtian) Malagasy taxon Majungasaurus crenatissimus, coupled with recent progress in morphometric analysis of theropod teeth, provide an opportunity to document dental morphology and quantitatively evaluate positional variation in an abelisaurid dentition. Majungasaurus possesses an unusually static dental formula of 4/17/17 and a relatively even pattern of tooth replacement. The teeth are brachydont, moderately heterodont, and exhibit fairly well-developed interdenticular sulci/caudae. The premaxillary teeth are significantly wider, more elongate, and less curved than those in the maxilla and dentary, whereas the dentary crowns are shorter and more ‘squat’ than those in the upper dentition. Knowledge gained from teeth preserved within jaws demonstrates that isolated crowns recovered from the same deposits in the Maevarano Formation of the Mahajanga Basin, and assigned to Majungasaurus by earlier workers, can be confidently referred to that species. Qualitative and quantitative differences are demonstrated between the teeth of Majungasaurus and those of the only other known non-avian theropod in the Maevarano assemblage, the noasaurid Masiakasaurus knopfleri. Documentation of the intra-individual and intraspecific variation in Majungasaurus tooth anatomy provides a standard against which other abelisaurids can be compared and provides a suite of dental features that potentially can be used to assist in evaluation of the phylogenetic position of Majungasaurus among abelisaurids and Abelisauridae among theropods (Smith, 2007).
Featured among the discoveries is an exquisite specimen (UA 8678) that preserves a virtually complete precaudal vertebral column, numerous costal elements, and portions of the skull and appendicular skeleton. This contribution represents the first description highlighting the postcranial axial skeleton of Majungasaurus. Owing to the completeness and quality of preservation, this specimen allows an examination of the serial transformation of features along the length of the axial skeleton, including a detailed analysis of postcranial pneumaticity in a nontetanuran theropod. Notable features of Majungasaurus include pneumatic cervical ribs with caudally bifurcate shafts and extensive pneumaticity of all postatlantal, precaudal vertebrae. Several postcranial features exhibited by Majungasaurus— including a well-developed cervical epipophysis, laterally expanded dorsal parapophysis, and sub-divided infradiapophyseal fossa in middle dorsal series—support previous phylogenetic studies placing it within Abelisauroidea and Abelisauridae. Majungasaurus (and abelisaurids generally) exhibit a robust cervical skeleton that features tightly interlocking cervical ribs, hyperossification of cervical rib shafts, and hypertrophied muscle attachment sites relative to other basal theropods. These features together highlight an axial core constructed to withstand high stresses, likely reflecting feeding adaptations for predation on large-bodied prey (O'Connor, 2007).
The appendicular skeleton of the abelisaurid theropod Majungasaurus crenatissimus (Deperet, 1896) Lavocat, 1955 is described for the first time. The available materials include an incomplete pectoral girdle and forelimb, along with the ilium and a nearly complete hind limb. These materials display a number of ceratosaur, abelisauroid, and abelisaurid synapomorphies, supporting the phylogenetic placement of Majungasaurus based previously on cranial anatomy. As in Ceratosaurus and Carnotaurus, the scapular blade is relatively wide and has a pronounced dorsal lip over the glenoid. The humerus is short and bears a globular head, but is more slender than in Carnotaurus. The ilium has a preacetabular hook, a strong supraacetabular crest, a notched posterior margin, and peg-and-socket articulations with both the pubis and ischium. Hind limb elements are proportionally stocky, as in some other abelisaurids. The femur lacks a trochanteric shelf, the tibia has a greatly enlarged cnemial crest, and the fibula bears a deep, posteriorly facing medial fossa. The abelisaurid astragalocalcaneum is described here in detail for the first time, and is more similar to that of tetanurans than to those of coelophysoids. Taken together, these materials illustrate that the appendicular skeleton of abelisaurids was specialized over the typical condition in basal theropods, particularly through the development of enlarged muscle attachment processes. The appendicular morphology of the abelisaurid theropod Majungasaurus crenatissimus is described. Abelisaurid appendicular materials have not been well documented, and several relatively complete specimens of this Late Cretaceous Malagasy theropod greatly clarify this region of the skeleton in these theropods. The forelimb of Majungasaurus is similar to those of other abelisaurids and includes a short, highly modified humerus. Numerous abelisaurid and abelisauroid synapomorphies are also found in the pelvis and hind limb. In addition, the pelvis and hind limb display a combination of features that strongly suggest a close affinity between abelisauroids and tetanurans. Functionally, the abelisaurid forelimb remains obscure and its interpretation must await more complete materials. The hind limb appears to show specializations for strong knee extensors, as well as unusual modifications of the metatarsus and pes that may have additional locomotor implications. (Carrano, 2007).
Recent collecting efforts (1993 to present) in the Upper Cretaceous (Maastrichtian) Maevarano Formation of northwestern Madagascar have yielded numerous specimens of the medium-sized abelisauroid theropod Majungasaurus crenatissimus (Deperet, 1896) Lavocat, 1955. Exquisite preservation, coupled with near-complete articulated or associated skeletons, has allowed an assessment of individual skeletal variation in this taxon, including a preliminary analysis of osteopathology in a non-tetanuran theropod. Importantly, Majungasaurus is the first abelisauroid theropod for which comprehensive pathology data are available. In a survey of 181 postcranial skeletal elements from a minimum of 21 individuals, abnormalities were identified in eight elements from a minimum of four individuals. These include a pedal phalanx with a mediolateral diaphyseal expansion of unknown etiology, a dorsal vertebra with a small exostosis, a caudal vertebra with probable idiopathic ossification of vertebral ligaments, and a pathologically truncated series of distal caudal vertebrae. The latter occurrence represents the first identified caudal truncation in a predatory dinosaur and only the second documented occurrence among non-avian dinosaurs (Farke and O'Connor, 2007).
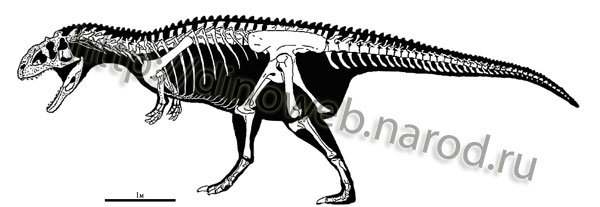
Illustrations: Majungasaurus crenatissimus skeleton Summary of phylogenetic, temporal, and biogeographic relationships of nonavian theropod genera from the Cretaceous of Gondwana. Taxa include most named genera recovered from Cretaceous deposits on Gondwanan landmasses (modified from Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998). Stratigraphically calibrated phylogeny of Abelisauridae (modified from Krause, Sampson, Carrano and O'Connor, 2007).
References: Deperet, 1896. Note on the sauropod and theropod dinosaurs from the Upper Cretaceous of Madagascar. Bulletin de la Societe Geologique de France, 3rd series, vol. 24 176-194.
Thevenin, 1907. Dinosauriens (Paleontologie de Madagascar IV). Annales de Paleontologie. 2, 121-136.
Lavocat, 1955. Etude des gisements de Dinosauriens de la region de Majunga (Madagascar). Travaux du Bureau Geologique. 69, 1-19.
Lavocat, 1955. Sur une portion de mandibule de Theropode provenant du Cretace superieur de Madagascar. Bull. Mus. Hist. Nat. Paris 27 256-259, 1 fig.
Lavocat, R., 1957. Sur les couches a dinosauriens de Madagascar. CCTA and Service Geologique de Madagascar, Comptes Rendus. Comites regionaux Centre, Est et Sud Conference de Tananarive, Avril 1957, Geology, Second Volume: 363-364.
Sues and Taquet, 1979. A pachycephalosaurid dinosaur from Madagascar and a Laurasia-Gondwanaland connection in the Cretaceous. Nature. 279:633-635.
Sues, H.D. 1980. A pachycephalosaurid dinosaur from the Upper Cretaceous of Madagascar and its paleobiogeographical implications. Journal of Paleontology 54:954–962.
Sampson, Krause, Dodson and Forster, 1996. The premaxilla of Majungasaurus (Dinosauria: Theropoda) with implications for Gondwanan Paleobiography. Journal of Vertebrate Paleontology. 16 (4): 601-605.
Sampson, Witmer, Forster, Krause, O'Connor, Dodson and Ravoavy, 1998. Predatory dinosaur remains from Madagascar: Implications for the Cretaceous biogeography of Gondwana. Science, 280: 1048-1051.
Rogers, R. R., D. W. Krause, and K. Curry Rogers. 2003. Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature 422: 515–518.
O'Connor, P. M. 2006. Pulmonary pneumaticity: an evaluation of softtissue influences on the postcranial skeleton and the reconstruction of pulmonary anatomy in archosaurs. Journal of Morphology 267: 1199–1226.
Krause, Sampson, Carrano and O'Connor, 2007. Overview of the history of discovery, taxonomy, phylogeny, and biogeography of Majungasaurus crenatissumus (Theropoda: Abelisauridae) form the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 1-20.
Rogers, Krause, Curry Rogers, Rasoamiaramanana and Rahanarisoa, 2007. Paleoenivronment and paleoecology of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 21-31.
Sampson and Witmer, 2007. Craniofacial anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 32-102.
Smith, 2007. Dental morphology and variation in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 103-126.
O'Connor, 2007. The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 127-162.
Carrano, 2007. The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 164-179.
Farke and O'Connor, 2007. Pathology in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. in Sampson and Krause (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. SVP Memoir 8. 180-184.
|  DINOWEB - палеозойские и мезозойские тетраподы
DINOWEB - палеозойские и мезозойские тетраподы